
Открытая нами феноменология кардинальной реорганизации архитектоники интерфазных ядер в онто- и филогенезе малярийных комаров и дрозофилы очевидно имеет общебиологическое значение. Основное затруднение в выявлении подобных закономерностей у других групп организмов связано с методическими ограничениями анализа топологии хромосомного аппарата в генеративной ткани, и прежде всего в яйцеклетке.
Подчеркну, что именно анализ генеративной тканевой системы позволил нам выявить наиболее существенные закономерности. Так, были обнаружены принципиальные различия архитектоники хромосом, с одной стороны, в генеративной сфере и соматических тканях, с другой - в самой генеративной ткани у разных (в том числе и очень близких) видов. Видоспецифичность проявляется в следующих показателях: 1) наличие - отсутствие связей хромосом с ядерной оболочкой и локализация связей на хромосомах; 2) морфология хромосомных участков прикрепления; 3) межвидовая разобщенность мест прикрепления гомеологичных хромосом на ядерной оболочке.
Другими словами, мы показали, что как в хромосомах, так и на мембране ядра существуют локальные, генетически детерминированные зоны, в которых обеспечивается жесткая связь хромосомы с мембраной. Хромосомы могут прикрепляться к мембране центромерными районами или интеркалярными локусами. В зонах прикрепления политенных хромосом к ядерной мембране могут быть пространственно разобщены как гомологи (асинаптическое прикрепление), так и плечи одной хромосомы по центромерным районам. Система взаимоотношений хромосом с ядерной мембраной облигатно инвариантна в пределах каждого вида малярийных комаров и четко различается между семью изученными видами. У межвидовых гибридов проявляются видоспецифичные особенности каждого из видов, причем значительная пространственная разобщенность зон прикреп-ления каждого из гомеологов в гибридных клетках свидетельствует о видоспецифичности координатных точек прикрепления на мем-бране.
Обнаруженный феномен хромосомной реорганизации нами рассматривается как явление мутационного ранга. Между тем ее нельзя отнести ни к одному из известных типов хромосомных мутаций, и мы оцениваем перестройку архитектуры интерфазного ядра как принципиально новый тип мутаций, названный нами системными мутациями. Здесь есть близость нашей трактовки с известными системными мутациями Рихарда Гольдшмидта, однако, как будет далее обсуждаться, эта аналогия не полная и лишь касается феноменологии "перетряски" интерфазного ядра.
Известный немецкий генетик Рихард Гольдшмидт - автор классических исследований по физиологической генетике и генетике пола, прошел парадоксальный путь в развитии своих эволюционных взглядов. В течение 20 лет он изучал микроэволюционные процессы в популяциях непарного шелкопряда. Сперва убежденный дарвинист, он пришел к заключению, противоречащему теории Дарвина:
формирование географических рас не ведет к видообразованию, естественный отбор не имеет конструктивного начала, а лишь устраняет неудачных мутантов. В начале 30-х гг. он радикально преобразовался в макромутациониста де-фризовского толка и в книге "Материальные основы эволюции" [Goldschmidt, 1940] аргументированно изложил свои взгляды на эволюцию. Мы еще будем касаться его эволюционных воззрений, здесь же заострим внимание на трактовке системных мутаций, которым он отводил ведущую роль в видообразовании. Гольдшмидт рассматривает вид как генетически закрытую систему, или, как он выражался, независимую реакционную систему (a single reaction system). Межвидовые изменения не могут быть вызваны изменением степени сложности или аддитивностью мелких мутаций - это полное преобразование исходного паттерна или реакционной системы в новую, которая затем может снова продуцировать внутривидовые вариации посредством микромутаций. Гольдшмидт называет этот тип генетических изменений системной мутацией и полагает, что в ее основе лежит преобразование хромосомного паттерна. Возникает новая, пространственно отличная упорядоченность хромосомного аппарата. Этот новый паттерн, видимо, формируется постепенно в серии последовательных этапов, которые могут проходить без видимого эффекта до тех пор, пока реорганизация хромосом не приведет к появлению новой стабильной системы.
Таким образом, Гольдшмидт полагал, что помимо известных микро- (генных) и макро- (хромосомных) мутаций должен существовать еще один тип - системные мутации, которые связаны с перетасовкой (scrambling) интимной хромосомной архитектуры, происходящей в редких случаях и предопределяющей удачное (или неудачное) событие, связанное с видообразованием [Goldschmidt, 1952].
Идея Гольдшмидта подверглась ожесточенной критике большинства эволюционистов 40-х - 60-х гг. (Э. Майр, Ф. Добржанский, Дж. Симпсон). В то же время ряд крупных биологов (А. Дальк, О. Шиндевольф, К. Уоддингтон) отнеслись к "системной мутации" с сочувствием, находя новые факты и теоретические подходы в се защиту.
Пожалуй, наиболее близок к нашему пониманию принципов системной реорганизации архитектоники генома генеративной сферы был бельгийский зоолог Альберт Дальк [Dalcq, 1949]. Его термин "онтомутация" был предложен для обозначения глубоких, резких, радикальных и вместе с тем жизнеспособных трансформаций, возникающих в цитоплазме яйцеклетки как морфогенетической системы. Дальк считал, что ранние стадии эмбриогенеза, а также главные морфологические параметры развивающегося организма детерминируются еще до оплодотворения - в ходе оогенеза. Как и Гольдшмидт, Дальк полагал, что, помимо реорганизации цитоплазмы, онтомутация связана с общим изменением всей ядерной системы. Однако, если системные мутации по Гольдшмидту - это единичные события и главной трудностью у "многообещающего урода" при этом является отсутствие партнера для спаривания, то для онтомутации Далька это затруднение снимается. Онтомутации вызываются резкими изменениями внешних факторов среды, которые одновременно влияют на всех самок в популяции в период созревания яйцеклеток. Воздействие среды на ооциты осуществляется в период их развития как непосредственно, так и за счет окружающих их питающих и фолликулярных клеток. Такие воздействия среды могут быть однонаправленными и многократными, вызывая накопление идентичных онтомутации в популяции [Dalcq, 1955, 1957 ].
Одновременно с Дальком сходную концепцию развивал французский биолог Альбер Вандель, считавший, что именно эмбриология, а не генетика стимулирует прогресс эволюционной теории:
зародышевые изменения, происходящие в яйце, а не генные мутации - суть эволюционных реорганизаций. Организация яйца (его общая архитектура) является основой для сопоставления видов и филогенетических групп [Vandel, 1948].
Таким образом, идея глубокой реорганизации хромосомного аппарата или всей ядерной системы в генеративной ткани организмов при видообразовании имеет серьезную историческую основу. При этом следует подчеркнуть, что ни Гольдшмидт, ни Дальк, ни Вандель или их сторонники не имели реальных доказательств правильности своих взглядов. Хотя Уоддингтон пытался обосновать реальность системных мутаций данными по лабораторным культурам клеток млекопитающих, где нередко происходили "перетряски" генома, сопровождавшиеся крупными морфологическими изменениями [Waddington, 1969], все же идея оставалась гипотезой вплоть до 1979 г., когда у малярийных комаров были выявлены именно такие реорганизации, которые с полным основанием можно было назвать системными мутациями [Стегний, 1979а].
Большинство хромосомных мутаций связано с изменением линейного расположения хромосомных блоков внутрихромосомного (инверсии) или межхромосомного (транслокации, центрические слияния) характера. Эти типы перестроек связаны не с количественными изменениями хромосомного материала, а лишь с его перераспределением в пределах одной хромосомы или всего набора. Другие хромосомные перестройки - типа дупликаций или делеций изменяют количество хромосомного материала. Есть основания считать, что все типы хромосомных мутаций, за исключением центических слияний, относятся к разряду необратимых событий, уникальных в смысле локализации хромосомных разрывов, возникающих при образовании этих перестроек. Уникальность хромосомных перестроек подчеркивает их принципиальное отличие от генных (аллельных) мутаций и особое значение в эволюционных событиях. Хромосомные перестройки относятся (по Гольдшмидту) к макро-мутациям, имеющим как внутривидовое значение (например инверсионный полиморфизм), так и видообразовательный смысл (фиксированные инверсии, транслокации, центрические слияния). Эволюционное значение хромосомных перестроек на примере инверсий мы рассмотрим в следующей главе.
Геномные мутации связаны с изменением количества хромосом, возникают в силу нарушений в аппарате веретена и бывают кратными (например полиплоидия) и не кратными (анеуплоидия). С этим типом мутаций в основном связывают видообразование (полиплодия), иногда внутривидовой полиморфизм (чаще анеуплоидия). По мнению В. Гранта [1991], подавляющее большинство папоротникообразных и покрытосеменных растений произошло за счет полиплоидии.
В чем же заключаются отличия принципиально новой хромосомной реорганизации, впервые обнаруженной нами у малярийных комаров [Стегний, 1979а] и названной системной мутацией [Стегний, 19876 ], от вышеприведенных общеизвестных типов мутаций? При системных мутациях не изменяется ни генный состав, ни линейная хромосомная структура, ни количество хромосом (или хромосомных участков). Таким образом, их нельзя отнести ни к одному из известных типов мутаций.
Системные мутации возникают в результате пространственной реорганизации интерфазных хромосом в ядре за счет изменения хромосомно-мембранных взаимоотношений. Другие особенности системных мутаций связаны с тем, что они являются видоспецифичными и не обнаруживают внутривидового или популяционного полиморфизма (при анализе большого числа особей у разных видов);
четко выявляются только в геномах генеративной ткани (в частности в трофоцитах), тогда как в изученных соматических тканях их визуализацию провести не удалось.
Происхождение системных мутаций сопряжено с реорганизацией хромоцентрального аппарата, которая заключается в появлении (или исчезновении) локусов прикрепления на хромосомах и на ядерной мембране и в динамичном изменении структуры хромоцентра (от локального до диффузного). Основную роль в прикреплении хромосом к мембране играет бетта-гетерохроматин, тяжи которого, в отличие от компактно-блочного альфа-гетерохроматина, внедряются в ядерную мембрану [Стегний, Шарахова, 1991 ]. У дрозофилы бетта-гетерохроматин редуплицируется при формировании политенной хромосомы и состоит в основном из повторов [Miklos, Cotsell, 1990 ], имеет, очевидно, происхождение из немобильных членов транспозиционных элементов типа Mdg-4 [Chantal et al., 1989]. Альфа-гетерохроматин состоит из сателлитной ДНК [Yamamoto et al., 1990], и для него характерна сильная недорепликация на политенных хромосомах. Гипотетически мы считаем, что наличие или отсутствие связи хромосомы с ядерной оболочкой определяется соответственно наличием или отсутствием бетта-гетерохроматина в соответствующей зоне. Так, оба плеча хромосомы 2 у Anopheles beklemishevi жестко прикреплены центромерными областями к ядерной мембране, причем хорошо видны тяжи бетта-гетерохроматина, внедряющиеся в оболочку. У других изученных видов малярийных комаров связи хромосомы 2 с ядерной мембраной не обнаружено, как и не отмечено наличия бетта-гетерохроматина в ее центромерной области. Имеются данные, что бетта-гетерохроматин может быть в центромерной зоне у одних видов (Drosophila uirilis, D. melanogaster) и отсутствовать у других (D. hydei) [Miklos, Cotsell, 1990]. Что касается структуры хромоцентра, то как у малярийных комаров, так и у дрозофилы существуют виды с сильным рассредоточением хромосом (и их плеч) по местам прикрепления к ядерной мембране (Anopheles messeae, Drosophila melanogaster). Другие виды демонстрируют более локальное распределение хромосом (Anopheles sacharovi, Drosophila sechellia) или же типичную хромоцентральную организацию (D. оrепа). Некоторые виды отличаются отсутствием контактов отдельных хромосом с оболочкой ядра (A. subalpinus, D. mauritiana, D. yakuba). В общем можно сказать, что хромосомальный аппарат - чрезвычайно динамичная структура и его "модификации" от локального до диффузного размещения на периферии ядра в составе ядерного матрикса имеют большое значение в формировании видоспецифичности взаиморасположения хромосом на основе хромосомно-мембранных связей. Возможные механизмы возникновения системных мутаций мы обсудим чуть ниже и теперь перейдем к оценке онтогенетических аспектов реализации системных мутаций.
Видоспецифичность организации хромосомного аппарата трофоцитов яичников ставит немаловажный вопрос: имеет ли эта структурная организованность отношение к функции питающих клеток - обеспечение накопления упорядоченной морфогенетической информации и запасных питательных веществ в цитоплазме яйцеклетки? Можно предположить, что имеет, причем элементы упорядоченности ооплазмы могут формироваться на основе топологической информации, которая свойственна геному каждой из питающих клеток в отдельности, и в то же время - на основе целостной системной информации, определяющейся взаиморасположением геномов всей группы трофоцитов по отношению друг к другу и к ооциту.
Изученный в плане групповой архитектоники геномов трофоцитов малярийный комар Anopheles messeae, как и другие виды, имеет 7 трофоцитов, расположенных на полюсе фолликула, противоположном локализации ооцита (рис. 3.1,6). При этом трофоциты формируют сферообразную систему, выпуклую по отношению к ооплазме. Три трофоцита, непосредственно прилегающих к ооплазме, отличаются более крупными размерами (и более крупными хромосомами). Четыре трофоцита, имеющих периферическое расположение, уступают размерами, при этом отношение диаметров самого мелкого ядра второй группы и самого большого ядра первой группы составляет около 1:2. Очевидно, что и степень политенизации хромосом трофоцитов также различна в данных группах клеток. В связи с этим можно с определенностью сказать о зна-чительно более высокой активности синтеза РНК в трофоцитах, прилегающих к ооплазме.
Учитывая, что топология хромосом всех трофоцитов вида идентична, что было доказательно представлено в главе 2, мы попытались ответить на вопрос: существует ли групповая упорядоченность хромосом всех семи трофоцитов по отношению друг к другу и к ооплазме? Методически это оказалось довольно трудно, так как необходимо было оценивать нераздавленный фолликул, в котором не нарушена организация трофоцитов. С другой стороны, наличие жестких хромосомно-мембранных связей хромосомы 3 и Х-хромосомы облегчало решение задачи, так как участки прикрепления хромосом к мембране являются хорошими маркерами суперпозиции отдельных трофоцитов по отношению друг к другу и к ооплазме. После удаления оболочки фолликула, состоящей из мелких клеток эпителия, с помощью стереомикроскопа можно было визуально оценивать взаиморасположение ядер трофоцитов по "торчащим" центромерным участкам хромосомы 3 (см. рис. 3.1,а), связанным с оболочкой ядер.
Первым, кто почувствовал видоспецифичную организацию генеративной ткани, был, по-видимому, известный цитолог конца прошлого века Оскар Гертвиг [1894]: "В то время как гистологические признаки ясно выражаются для нас в продуктах дифференцировки тканей, видовые свойства клеток ускользают от нашего прямого наблюдения, так как мы в настоящее время не можем проникнуть в область явлений мицеллярной организации клеток. Только научные выводы общего характера вынуждают нас допустить, что существует организация, лежащая вне постижения нашими чувствами. Затронутый здесь пункт лучше всего разъясняется на примере половых клеток". Гертвиг полагал, что как женские, так и мужские половые клетки одного вида организмов практически тождественны по своим существенным свойствам. В то же время половые клетки различных видов различаются по "формуле состава" и располагаются соответственно группировке видов животных и растений в естественной системе. Гертвиг представил клетку в качестве единицы меры для исследования органического мира. "Соображения такого рода привели нас к понятию о видовой клетке, то есть клетке, в тончайшей (мицеллярной) организации которой содержатся существенные свойства вида в качестве определяющих частей, выраженных в системе клеточных свойств. Сколько видов в растительном и животном царстве, столько же можно различить и видовых клеток. Они являются представителями видов, существенные признаки которых сведены в них к простейшей формуле".
Все известные до настоящего времени мутации подразделялись на три группы: генные (точковые), хромосомные и геномные. Преобладающее большинство генных мутаций относится к разряду аллельных вариантов того или иного гена и являются обратимыми. Этой группе, по нашему мнению, наиболее соответствует термин "геновариация", предложенный С. С. Четвериковым в 1926 г. и, к сожалению, вышедший из употребления. Другие генные мутации (например микроделеции) могут являться необратимыми. По определению Гольдшмидта генные мутации относятся к уровню микромутаций (аналогия с термином "микроэволюция" Ю. А. Филипченко), имеющих отношение к внутривидовой генетической адаптации, но не к видообразованию.
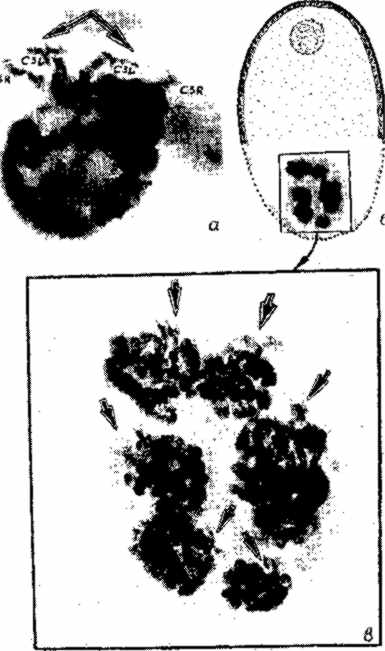 |
| Рис. 3.1. Взаиморасположение ооцита, ооплазмы и хромосомно-мембранных контактов группы трофоцитов у малярийного комара Anoplieles niesseae. а - ядро трофоцита, СЗR, C3L - центромерные участки хромосомы 3; б - общий вид фолликула; в - увеличенный снимок группы трофоцитов. Стрелками показаны зоны прикрепления хромосомы 3 к ядерной оболочке. |
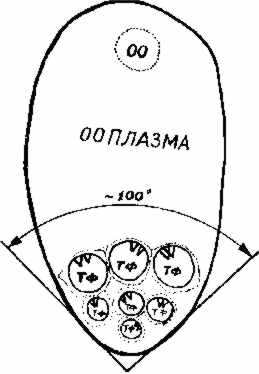 |
Рис. 3.2. Схема взаиморасположения ооцита (00), ооплазмы и трофоцитов (Тф) малярийного комара Anopheles messeae. На трофоцитах показаны места прикрепления хромосомы 3. Угол разброса точек прикрепления по отношению к ооциту и ооплазме составляет около 100° |
Анализ большого числа фолликулов, удачно ориентированных боковой областью перпендикулярно оси объектива, показал, что центромерные участки хромосомы 3, связанные с ядерной мембраной, направлены в сторону ооплазмы у всех 7 трофоцитов фолликула (б, б). При этом, если мысленно провести осевую линию вдоль фолликула, то направленность прикрепленных участков хромосомы 3 может колебаться в пределах 100° к плоскости ооплазмы (рис. 3.2). Учитывая сферичность организа-ции группы семи трофоцитов. можно предположить, что эти отклонения связаны с ошибкой визуального наблюдения. XL-плечо Х-хромосомы у A. messeae прикреплено к ядерной оболочке боковой областью (см. рис. 2.4,а). Об ориентации XL-плеча в ядрах 7 трофоцитов можно было судить по редким удачно расположенным группам трофоцитов.
Дело в том, что при просмотре ядер в дорзовентральной (боковой) плоскости, когда возможна четкая идентификация прикрепленных к оболочке плеч 3R и 3L, хромосому XL обнаружить не удавалось. Если же (в редких случаях) наблюдение велось с "полюса" (торцевой части фолликула), то XL-плечо оказывалось видимым, тогда как хромосома 3 исчезала из поля зрения. Поэтому мы предположили, что плоскость расположения XL-плеча перпендикулярна плоскости центромерной области хромосомы 3, связанной с оболочкой ядра.
Таким образом, можно сделать вывод о коориентации хромосомных структур группы трофоцитов по отношению к ооплазме и собственно ооциту. Открытие этого явления позволяет гипотетически оценить его роль в формировании оогенетической сегрегации.
Еще недавно доминировало мнение, что морфогенез - формирование зародыша из оплодотворенной яйцеклетки - представляет собой образование сложных пространственно упорядоченных форм из изотропной цитоплазмы зиготы. Вместе с тем уже в начале XX в. классические работы Конклина продемонстрировали явление ооплазматической сегрегации, т. с. структурной упорядоченности цитоплазмы зрелой и оплодотворенной яйцеклетки с полной детерминацией всех исходных стадий морфогенеза. По сути, современные молекулярно-генетические данные окончательно убеждают в строгой ооплазматической сегрегации.
В Базельском университете у дрозофилы были выделены и клонированы гены гомеозисных мутаций [Hafen et al., 1984 ]. С помощью РНК-ДНК гибридизации было показано, что РНК-копии генов, ответственных за формирование отдельных сегментов организма, образовывали четкие скопления в совершенно определенных зонах цитоплазмы ооцита, т. е. наблюдалась строгая структурированность цитоплазмы без каких-либо клеточных перегородок. Эти данные свидетельствуют о фундаментальном принципе организации яйца - регионализации, т. е. распределении информационных макромолекул по определенным участкам цитоплазмы, причем локализация этих участков тесно коррелирует с планом дробления или первых этапов эмбриогенеза.
Следует подчеркнуть, что хотя в современной эмбриологии оогенетическая сегрегация - вполне доказанное явление [Айзенштадт, 1984; Рэфф, Кофмен, 1986 ], механизмы ее становления остаются совершенно "темной" проблемой, от решения которой зависит выяснение механизмов дальнейших этапов онтогенеза.
Попытаемся найти подходы к объяснению феномена оогенетической сегрегации с позиций выявленных нами топологических закономерностей в организации геномов трофоцитов. Напомним, что именно геномы трофоцитов ответственны за развитие ооплазмы у организмов с алиментарным типом оогенсза, к которым относятся малярийные комары и дрозофила. Связь ооцита с каждым из трофоцитов осуществляется цитоплазматичсскими мостиками или кольцевыми каналами [King, 1970].
Формирование яйцевой клетки, как и в целом проблема онтогенеза, рассматривается в современной биологии с позиций трех основных направлений: 1) генетическая теория развития (Дэвидсон); 2) теория физиологических градиентов (Чайлд); 3) теория поля (Гурвич, Кольцов). Первые два направления активно обсуждаются во всех современных сводках по проблемам онтогенеза, поэтому мы не будем на них заострять внимание [Современные ..., 1977; Нейфах, Тимофеева, 1977; Айзенштадт, 1984; Газарян, Белоусов, 1983 ]. Отметим лишь, что эти теории, продуктивные во многих отношениях, бессильны объяснить механизмы становления первичных этапов оогенетической сегрегации. Методология "полевого" подхода базируется на идее "биологического поля", развитой в 30-х гг. гистологом и эмбриологом А. Г. Гурвичем и незаслуженно забытой в настоящее время. Первоначально Гурвич мыслил "поле" как некий материальный фактор, однако впоследствии склонился к тому, что "сама идея поля сможет быть со временем облечена и в язык физики". Стержневым моментом в его концепции был поиск таких инвариантных законов, которые могли бы работать в условиях "неудержимости эмбриогенеза", при непрерывной смене субстрата - "в потоке". "Сначала инвариантность любой ценой, а потом происхождение и идентификация инвариантного начала". Гурвич был противником того, чтобы общую теорию наследствен-ности оценивать только через менделевские принципы. "Формальная генетика оперирует отдельными признаками (доступными для гиб-ридологического анализа) различной природы: химические, цито-логические, макроморфологические. Считается, что организм может быть поделен на признаки (без остатка). Это недопустимо. При-знаки - это конечный результат процессов развития, но анализ самих процессов генетика не считает своей задачей... Мы... при-ходим к логической необходимости признать истинно реальным объектом наследственности сам процесс осуществления типичного хода эмбрионального развития." На основе экспериментов Гурвич показал, что в цитоплазме имеются упорядоченные группировки тех или иных молекул, которые он назвал неравновесными моле-кулярными констелляциями. Существование подобных систем он объяснил наличием неких ориентирующих факторов, или поля.
Источником поля Гурвич считал молекулы ядерного хроматина в момент их синтеза (образования комплекса ДНК - белок), при этом поле ядра существует постоянно, преемственно передаваясь через молекулы ДНП. Суммарный источник поля сосредоточен в центре ядра. Наиболее равновесным положением ядра является геометрический центр клетки. Любая "скошенная" конфигурация клетки будет неравновесной. Каждый источник создает вокруг себя пространственно анизотропное поле, т. е. векторы поля неодинаковы по трем направлениям пространства. Это можно выразить эллипсоидом с тремя неравными осями, описанным вокруг источника поля. Конфигурация эллипсоида анизотропии принимается за видовую постоянную и является в теории поля единственно наследственно преформируемым началом, из которого эпигенетическим путем выводится разнообразие органических форм [Гурвич, 1944].
Этот последний постулат является для нас отправным пунктом для обоснования механизмов становления оогенетической сегрегации на основе видоспецифичной пространственной организации хромосомного аппарата трофоцитов. Прежде чем перейти к формулировке нашего представления, отметим, что физический смысл "полю Гурвича" придал Н. К. Кольцов [1936 ], описавший развитие яйца в терминах электрического силового поля. Оно "создается благодаря выводу из ядра в ооплазму веществ с различными электрическими потенциалами и оно ориентировано согласно общему плану строени ооцита, но детализирующего и закрепляющего этот план. Еще до созревания этот план в существенных чертах намечен, преформирован". В 1930 г. Спэк [Кольцов, 1936] обнаружил в яйце Nereis катафорезные токи, возникающие за счет разницы потенциалов и обеспечивающие перемещение веществ в ооплазме.
На современном уровне это убедительно подтвердили Вудруфф и Телфер [Woodruff," Telfer, 1973] на бабочке цекропии. Переход рРНК из трофоцитов в ооцит осуществляется в составе РНП-частиц (типа рибосом) не посредством градиента концентрации, а за счет разницы потенциалов ооцита и трофоцитов. С помощью микроздектродов в фолликулы был инъецирован меченный флуорохромом белок. Оказалось, что белок может проходить по цитоплазматическим мостикам от трофоцита к ооциту, но не наоборот. Авторы с помощью внешнего напряжения (с током 5x10-8А) смогли изменить полярность, и меченый глобулин стал переходить из ооцита в трофоциты. Измерение потенциала в трофоцитах дало -40 мВ, в ооцитах -30 мВ. Таким образом, разность потенциалов в 10 мВ и является основным условием перехода рибонуклеотидов из трофоцитов в ооцит. В течение нескольких дней содержание РНК в ооцитах цекропии увеличивается в 5 раз (от 0,5 мкг до 2,5 мкг), а в трофоцитах уровень РНК сохраняется постоянным (0,5 мкг).
Таким образом, электрокинетические принципы становления ооплазмы на основе общей теории поля Гурвича могут быть убедительно, с нашей точки зрения, соотнесены с выявленными нами системными мутациями. Гипотетически можно принять следующие постулаты.
Распространяя эту модель на последующие этапы онтогенеза, можно предполагать, что при морфогенезе происходит многократная реорганизация архитектоники интерфазных ядер, регуляторно (посредством модифицированных полей) обеспечивающая морфофункциональные различия отдельных клеточных систем или тканей. Пространственная реорганизация структуры интерфазного ядра происходит в инициальной (стволовой) клетке, и в пределах производной от нее клеточной системы или ткани ядра структурно идентичны (в норме), что определяет специфическую активность геномов, входящих в одну систему.
Имеются сведения, что биоэлектрическая активность клеток в онтогенезе существенно модифицируется и, по-видимому, играет определяющую роль в регуляции процессов морфогенеза [Шахбазов, 1970; Зубарев и др., 1974]. Биоэлектрогенез клеток обусловливается электрическими взаимодействиями между высокоупорядоченными молекулярными и надмолекулярными структурами, и прежде всего ядрышком и доменами хроматина. Пространственная организация этих систем во взаимодействии с ядерной мембраной предопределяет морфофункциональные особенности того или иного клеточного типа [Bourgeois, Hubert, 1988; Manuelidis, Borden, 1988 ]. Вместе с тем эта упорядоченность имеет динамическую основу. Показано, что ядрышки и домены хроматина испытывают криволинейные флуктуации и определяют ядерное вращение, которое происходит скачкообразно с периодами стационарности и изменением направления [De Boni, 1988].
Структурные модификации хроматина (конденсация - деконденсация) хорошо описываются в терминах электростатических взаимодействий [Clark, Kimura, 1990]. Экзогенные воздействия (гормональные, температурные) могут существенно влиять на электрокинетические характеристики хроматина, ядрышек и целых ядер [Шахбазов и др., 1986; Шахбазов, 1989]. Показано также, что внешние электромагнитные сигналы могут значительно стимулировать синтез РНК [Goodmen, Henderson, 1987 ], как и повторное прохождение определенных стадий морфогенеза при регенерации [Мелкумян и др., 1975].
Таким образом, учитывая все возрастающий интерес к клеточному биоэлектрогенезу, мы считаем, что поиск закономерностей электростатических взаимодействий клеток на основе архитектоники хромосомного аппарата будет перспективным в плане разработки подходов к решению кардинальных проблем оогенетической сегрегации, клеточной дифференцировки и морфогенеза. Мы убеждены, что пресловутый "гомункулус", или проформа организма, реализован именно в видоспецифичной топологии ядер клеток генеративной ткани (трофоцитов и ооцитов).
В заключение кратко рассмотрим возможные факторы, приводящие к возникновению системных мутаций. Поскольку происхождение системных мутаций определяется, прежде всего, реорганизацией хромоцентрального аппарата и формированием связей хромосом с ядерной оболочкой, то, видимо, следует обратить внимание на такие факторы, которые детерминируют модификации гетерохроматина и хромосом в целом в генеративных клетках. Анализ данных подобного рода привел нас к заключению, что жесткий инбридинг в сочетании с экстремальным температурным воздействием в некоторые периоды онтогенеза может способствовать появлению системных мутаций. Жесткий инбридинг представляет собой еще недостаточно ясное явление в плане генетических эффектов. Помимо известных явлений гомозиготизации аллельных и хромосомных вариантов, с ним связаны такие процессы, как резкое усиление активности мобильных элементов и мутабильности [Гвоздев, Кайданов, 1986], и, что нас более интересует, структурные преобразования в хромосомном аппарате генеративной ткани.
Так, было показано [Bier, 1958], что в трофоцитах мухи Calliphora erythrocephala в результате жесткого инбридинга в сочетании с низкотемпературным воздействием (культивирование при +16°С) наблюдаются существенные модификации хромосомного аппарата. Если в обычных лабораториях или диких популяциях хромосомы трофоцитов редко образуют политению и ядро имеет ретикулярную структуру хроматина, то при сестринском инбридинге к седьмому поколению в фолликулах начинают накапливаться трофоциты с политенными хромосомами. Количество таких трофоцитов и степень политенизации увеличивается с каждым поколением, и к 12-15-му поколениям их частота достигает 30-40 % от числа всех питающих клеток. Если бы этот эффект определялся переходом в гомозиготное состояние определенного гена (или группы), то все фолликулы яичника должны были бы сходно изменить хромосомную структуру трофоцитов с какого-то определенного поколения инбридинга. Тем не менее реальная картина показывает, что в ходе инбридинга постепенно нарастает дестабилизация нормального хромосомного паттерна, как и растет число дефектных яиц, из которых не развиваются эмбрионы.
Интерес представляет также работа, где обнаружено влияние инбридинга на возникновение модификаций гетерохроматиновых узелковых районов в мейоцитах кукурузы. Через 2-3 поколения изменяется число и размер узелковых районов и вместо обычного компактного гетерохроматина возникают хромомерно-диффузные образования [Похмельных, Шумный, 1984].
Признавая возможное значение инбридинга и температурных воздействий для образования системных мутаций, необходимо учитывать, что для экспериментов важно подбирать виды с подходящей структурой генома генеративной ткани. Речь идет о видах с так называемыми эволюционно лабильными геномами.