
В период бурного расцвета популяционной генетики (30-е - 70-е гг.) были созданы биологическая концепция вида и так называемая синтетическая теория эволюции (СТЭ), являющиеся по сути апофеозом дарвиновского понимания эволюционных преобразований. Биологическая концепция вида подвергала острой критике (в крайних позициях - вообще отвергала) традиционную систематику за формальный типологический подход к проблемам признака и таксономии [Майр, 1971]. В то же время синтетическая теория эволюции, базируясь на популяционно-генетическом мышлении, пыталась убежденно доказывать, что суть видообразования (как и образование таксонов более высокого ранга) есть плавное (градуальное) превращение популяции исходного вида в некоторую новую видовую популяцию, контролируемое естественным отбором [Майр, 1968; Dobzhansky, 1971].
Между тем к концу 70-х гг. наметился кризис популяционно-генетического подхода к эволюционным проблемам, базирующегося на редукционной методологии [Левонтин, 1978; Wright, 1980]. Десятилетие 80-х гг. прошло под знаком сомнения и переосмысления как принципов систематики, опирающихся на биологическую концепцию вида, так и основных положений синтетической теории эволюции. Доминирующим концепциям противопоставлялись идеи, которые базировались на возрождении типологического мышления в систематике [Мейен, 1984] и макромутационистском толковании проблемы видообразования (идеи С. Коржинского, Г. Де Фриза, Р. Гольдшмидта) [Altukhov, 1982; Стегний, 1982, 1983а; Тахтаджян, 1983 ].
Ниже представлен критический анализ ряда положений синтетической теории эволюции и, что более важно, впервые сделана попытка оценить значение системных мутаций с точки зрения видообразовательных событий и с позиций филогенетической систематики.
Современная систематика характеризуется, с одной стороны, развитием традиционного морфометрического анализа и, с другой - внедрением новых методов: хромосомного, биохимического, гибридологического и других. Привлечение этих методов способствует прогрессу в выяснении системы и в реконструкции филогении отдельных "трудных" таксонов и в то же время создает серьезные проблемы, связанные с понятием "таксономический признак", а именно с его значимостью при различных методических подходах.
Основные задачи систематики, по-видимому, наиболее точно определил А. А. Любищев [1971 ]: 1) дискриминация - четкое разграничение соседних таксонов; 2) идентификация - вполне надежное определение каждого экземпляра на любой стадии развития; 3) систематизация - приведение всех таксонов в естественную систему, выявление истинного порядка всего многообразия организмов. Решение этих задач зачастую бывает затруднено в связи с существованием сложных ситуаций в отдельных таксонах. Так, например, применение обычного морфометрического анализа не всегда дает возможность дискриминировать формы, которые по биологическим показателям ведут себя как "хорошие" виды. Эта ситуация, особенно часто встречающаяся у насекомых, известная как проблема видов-двойников, очень наглядно демонстрирует ограниченность морфометрических приемов. Привлечение гибридологического, хромосомного, биохимического методов обычно в этих случаях способствует четкому разграничению видов, причем в качестве таксономических здесь выступают такие видоспецифичные признаки, как структура (или количество) хромосом, электрофоретическая подвижность белков (ферментов) и другие. Однако сразу же подчеркнем, что эти таксономические признаки, скрытые от взгляда обычного систематика-морфолога, отнюдь не универсальны и каждый из последних методов имеет такие же ограничения (если не большие), как и морфометрический.
Попытаемся далее наметить пути разрешения проблем, возникающих при анализе трудных в систематическом отношении таксонов. В связи с тем что интересы автора связаны с отрядом Diptera, в работе в основном использованы данные по этим насекомым, тем не менее излагаемые принципы могут быть применены для самых разнообразных групп.
Как уже отмечалось, использование морфометрического метода при таксономическом анализе Diptera иногда сопряжено со значительными трудностями. Часто метод "работает" лишь на одной из стадий онтогенеза, а в некоторых случаях и вообще "не работает".
Например, в близкородственном комплексе Anopheles maculipennis (Culicidae) только различия в структуре и окраске яиц позволяют надежно разграничивать биологически дискретные формы, личиночные и имагинальные признаки которых практически идентичны [Гуцевич и др., 1970]. Аналогичная ситуация характерна и для многочисленных представителей Simuliidae [Чубарева, Петрова, 1979] и Chironomidae [Keyl, Keyl, 1959 ].
Целесообразность применения в таких случаях кариологического анализа определяется тем, что морфологические особенности хромосомного аппарата, по-видимому, всегда носят видоспецифический характер. Это связано с системной реорганизацией кариотипа при видообразовании. Следует отметить, что универсального хромосомного признака, имеющего таксономическое значение, вероятно, не существует. В одних случаях имеются межвидовые различия в числе и морфологии митотических или мейотических хромосом, как Drosophilidae [Stone et al., I960; Wasserman, 1963] или Tabanidae [Boyes, Wilkes, 1972], в других отмечается высокий консерватизм этих параметров. Так, у Culicidae [Kitzmiller, 1976] и Simuliidae [Чубарева, Петрова, 1979] хромосомы зачастую не дают систематикам видоспецифичной информации. Применяемые в таких случаях сложные морфометрические методы также не всегда успешны [Кабанова, Карташова, 1972 ]. Тем не менее крупные таксономические группы (подроды, роды, семейства) имеют обычно существенные различия в числе и морфологии метафазных хромосом. Можно предполагать, что перспективность использования этих хромосом для таксономических целей увеличится с применением методов их дифференциальной окраски [Morata, Rai, 1977].
Анализ гигантских (политенных) хромосом значительно эффективнее метафазного анализа и успешно используется в систематике семейств Chironomidae, Culicidae, Drosophilidae, Simuliidae и некоторых других. У многих Diptera политенные хромосомы отмечены в разных тканях у личинок (слюнные железы, кишечник, мальпигиевы сосуды) и имаго (мальпигиевы сосуды, питающие клетки яичников). Обычно структура и морфология дисков политенных хромосом имеют видоспецифичный характер, а межвидовые различия связаны с легко распознаваемыми фиксированными перестройками блоков хромосом (типа инверсий, дупликаций, делеций), изменением локализации центромерных участков, количества гетерохроматина и числа ядрышковых организаторов. Различия более высоких таксонов определяются, кроме перечисленных, сложными реорганизациями хромосом (типа транслокаций). Благодаря анализу политенных хромосом проведена ревизия и уточнена систематика в семействах Simuliidae [Чубарева, Петрова, 1979; Dimbar, 1966 ], Chironomidae [Keyl, 1962 ], Culicidae [Coluzzi et al., 1979; Стегний, 1981 ], Drosophilidae [Sturtevant, Dobzhansky, 1936; Stone et al., I960; Wasserman, 1963; Clauton et al., 1972]. Причем во всех этих группах были описаны новые виды, как правило, почти (или полностью) не различимые по внешней морфологии.
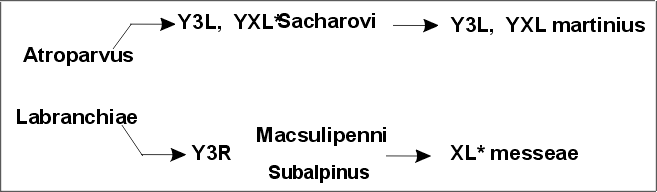
Цитогенетическое исследование палеарктического комплекса Anopheles maculipennis (Culicidae) способствовало четкой дискриминации форм по структуре политенных хромосом [Frizzi, 1947;
Стегний, 1981]. В настоящее время не вызывает сомнений видовой статус каждой из восьми репродуктивно изолированных форм комплекса [Стегний, 1980а]. На территории Северной Евразии выявлен новый вид Anopheles beklemishevi [Стегний, Кабанова, 1976; Stegnii, Kabanova, 1978], отличающийся от остальных видов фиксированными перестройками во всех хромосомах. Кроме того, благодаря хромосомному и гибридологическому анализу восстановлена валидность A. martinius, описанного Н. Шингаревым в 1926 г. [Стегний, 1980а]. Этот вид, обитающий в Средней Азии, отличается от A. sacharovi фиксированными хромосомными инверсиями [Стегний, 1976] и репродуктивно изолирован от последнего [Стегний, 1980а].
Большинство видов палеарктического комплекса maculipennis четко различаются хромосомными перестройками, и сравнительные фотокарты политенных хромосом могут служить своеобразным определителем этой подгруппы [Стегний, 1980а]. В то же время в этом комплексе имеются две группы видов: A.atroparvus - A.labranchiae и A.maculipennis - A. subalpinus, внутри которых структура политенных хромосом идентична. Между тем гибридологический анализ показал, что виды репродуктивно изолированы и, следовательно, имеют такую же самостоятельность, как и остальные палеарктические виды [Frizzi, De Carii, 1954]. Однако линейная структура политенных хромосом в данной ситуации не может служить таксономическим признаком.
| Вид | МДГ-3 | Г-6-ФДГ | КДГ | ОТ | МЭ |
| Anopheles atroparvs | 1,00 | 1,00 | 1,00 | 1,00 | 1,00 |
| A.maculipennis | 0,91 | 0,98 | 1,00 | 1,00 | 1,00 |
| A.messeae | 0,91 | 1.00 | 1,00 | 1,00 | 0,96 |
| A.sacharovi | 0,91 | 0,98 | 0.96 | 1,00 | 0,96 |
| A.martinius | - | 0,98 | - | - | - |
| A.beklemishevi | 0,85 | 0.96 | 0,92 | 0,72 | - |
Оценивая метод электрофореза белков для целей систематики, отметим один существенный его недостаток - отсутствие автономности. Без связи с другими таксономическими методами он вряд может применяться. Это связано с возможностью независимого сходства (электрофоретической идентичности) отдельных белков у неродственных таксонов. В то же время этот метод можно рекомендовать при анализе близкородственных форм (видов-двойников), как опыт показывает его эффективность в таких случаях, особенно если исследуются одновременно 5-10 белков.
Выявление филогенетических связей всегда сопряжено со многими проблемами, и в первую очередь, с возможностью полифилетического происхождения сравниваемых таксонов и с параллельным или конвергентным развитием отдельных таксономических признаков. Степень риска при выборе признаков, подходящих для выяснения системы и филогении видовых комплексов, определяется, вероятно, уровнем их внутривидового консерватизма (видоспецифичности) и характером межвидовых взаимоотношений (на уровне разнообразных биологических показателей).
Мы полагаем, что в основе филогенетических построений должен лежать комплексный методический подход, значительно повышающий надежность выводов, поскольку филогенез как система родственных отношений, очевидно, строится на многообразии биологических показателей. Проиллюстрируем это на примере выяснения филогенетических связей.
Структурные особенности политенных хромосом для филогенетических исследований впервые применили Стёртевант и Добржанский [Sturtevant, Dobzhansky, 1936], затем этот метод широко использовался и оказался чрезвычайно эффективным при анализе разнообразных таксономических групп Drosophilidae [Wesserman, 1963; Clauton et al., 1972], Chironomidae [Keyl, 1962], Culicidae [Coluzzi et al., 1979] и других. В основу метода положено допущение, что любая хромосомная инверсия уникальна по локализации точек разрывов на хромосоме и монофилетична по происхождению [White, 1954]. Таким образом, если три вида различаются заходящими инверсиями по какой-либо хромосоме, то можно провести реконструкцию связей этих видов и точно установить промежуточное звено в данной системе. Если же виды различаются заходящими инверсиями и по другим хромосомам и реконструкция межвидовых связей по каждой хромосоме совпадает, то значительно увеличивается вероятность того, что данная филогенетическая система естественна. Базируясь на теории монофилетического происхождения локальных инверсионных перестроек, можно заключить, что если несколько видов обладают идентичной структурной перестройкой по какой-либо хромосоме, то эта структура является филогенетически более древней по сравнению с любой другой уникальной инверсией. Такое положение согласуется с принципами филогении, разработанными Геннигом [Hennig, 1966], и позволяет оценить направление эволюции не только отдельных хромосом, но и целых видовых кариотипов.
Мы показали, что у малярийных комаров хромосомный тип atroparvus - labranchiae является филогенетически исходным для палеарктической группы maculipennis [Стегний, 1979б]. От этого хромосомного типа берут начало две филогенетические линии:
В схему не включен A.beklemishevi, поскольку его статус в Палеарктике относительно независим и он непосредственно связан с неарктическим хромосомным типом A.earlei [Стегний, Кобанова, 1976].
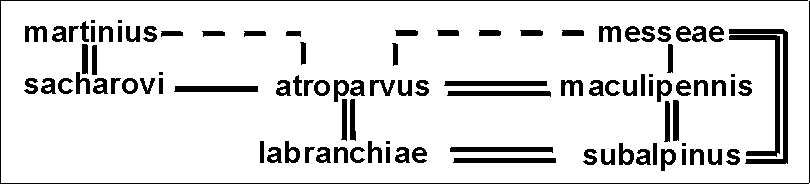
Корректность данной схемы видообразования в палеарктическом комплексе maculipennis, основанной на межвидовых хромосомных связях, подтверждается данными по лабораторной гибридизации видов этого комплекса. Показано, что существуют большие различия в степени выраженности репродуктивной изоляции между отдельными видами. На основе этих результатов была создана схема связей видов, отражающая степень близости их геномов [Стегний, 1980а]:
Примечание. Двойные, одинарные и пунктирные линии означают соответственно близкое (гибридные самки способны к возвратному скрещиванию, самцы стерильны), среднее (гибридные самки и самцы жизнеспособны, но стерильны) и дальнее родство (гибель гибридов на эмбриональных или личиночных стадиях); отсутствие линий между некоторыми видами означает, что скрещивание не проводилось.
Сопоставляя эти две схемы, построенные независимо (по разным критериям), можно видеть, что противоречия между ними нет. Хромосомная схема лишь более детально выявляет филетические линии и демонстрирует направление эволюционного процесса. Сравнительный анализ политенных хромосом и репродуктивных отношений палеарктических видов и неарктической группы maculipennis (более древней) однозначно показывает, что генератором видообразования в Палеарктике являлся один из видов группы A.atroparvus - A.labranchiae [Стегний, 1980а, 1981]. Относительная изоляция в Палеарктике A.beklemishevi, показанная на основе хромосомного анализа [Стегний, Кабанова, 1976], хорошо подтверждается данными по ферментным системам (см. таблицу). A.beklemishevi отличается от других изученных видов по всем четырем мономорфным локусам.
Остается неясным, какова роль каждого из четырех гомосеквентных видов в видообразовании, поэтому полная детализация филогенетических связей в палеарктическом комплексе maculipennis пока не проведена.
В целом же можно отметить, что только благодаря комплексному методическому подходу удалось получить многообразную информацию о системе и филогении изучаемой нами группы видов-двойников, и напомнить, что с точки зрения систематика-морфолога эта группа является практически безликой, а формы, в нее входящие, заслуживают лишь подвидового статуса.
Касаясь принципов "взвешивания" признаков, на основе которых устанавливаются систематические связи, следует отметить, что корреляция критериев сходства - различия и родства - неродства является для нас аксиомой, по крайней мере, для тех примеров, которыми мы оперируем. Принципы филогенетической систематики Геннига [Hennig, 1966] наиболее адекватно отражают дивергентный характер эволюции. Особенно ярко они проявляются на уровне "низших" систематических категорий, в частности группировок видов, родство которых легко доказать прямыми методами, используя критерий скрещиваемости. Для подобных видов, дискретность которых определяется репродуктивной изоляцией, важно найти признаки, наиболее полно отражающие уникальность генетической системы вида, т. е. генеалогические признаки. Виды малярийных комаров являются сходными по внешней морфологии имаго и личинок, хотя назвать их видами-двойниками в строгом смысле нельзя, так как они все же четко различаются по структуре и окраске яиц.
Уникальность генетической системы вида есть элементарное типологическое качество, отражающее таксономическую дискретность. Уникальность очевидно связана с инвариантной частью видового генома, а ее проявление реализуется в самых разных показателях инвариантности (внешняя морфология, хромосомы, ферменты и др.). Причем практически в одних случаях видовая дискриминация может осуществляться посредством "работы" многих показателей, в других - одного-двух. В этом смысле использование ряда относительно независимых параметров увеличивает, с одной стороны, надежность диагностики, с другой - вероятность быстрого нахождения удачного показателя. Наиболее значимыми для таксономии признаками представляются генетически детерминированные, не имеющие средовых искажений. Сюда относятся, например, хромосомные перестройки, ферментные локусы.
Особое значение в связи с этим имеют системные мутации, которые, как мы показали, надежно дискриминируют даже гомосеквентные (хромосомно-двойниковые) виды. Можно предположить, что системные мутации являются базовыми таксономическими признаками видового ранга. Мы реально оцениваем возражения или сомнения по поводу их универсальности по отношению к любому виду, поскольку в связи с методическими ограничениями при анализе той или иной группы организмов существует и проблема выявления системных мутаций. Однако, учитывая значение видоспецифичной архитектоники генома генеративной ткани для развития яйца, a priori можно полагать, что данный показатель полностью отражает специфику вида.
В заключение следует отметить, что традиционный морфологический анализ в систематике играет и в настоящее время ведущую роль. Ни один из современных методов не "работает" без предварительного анализа таксона. Углубляя и уточняя систему и филогению той или иной группы с помощью хромосомного, гибридологического или биохимического методов, мы, тем не менее, всегда за основу берем морфологическое описание данной группы.
Хромосомные механизмы видообразования и генетической адаптации видовых популяций представляют одну из актуальных проблем эволюционно-генетической теории. Многие вопросы хромосомной организации видов решены благодаря анализу многочисленных групп двукрылых насекомых, среди которых особо выделяются род Drosophila. В настоящем разделе предполагается рассмотреть ряд дискуссионных, по мнению автора, положений, связаных с сущностью хромосомных инверсий, эволюционной ролью инверсионного полиморфизма и хромосомными реорганизациями при видообразовании. Характерной чертой двукрылых насекомых является исключительная роль хромосомных инверсий в эволюции видовых кариотипов. Трудно переоценить значение инверсионного полиморфизма в генетической адаптации многих представителей Diptera. Помимо этого, филогенетические хромосомные связи близкородственных видов зачастую определяются фиксированными инверсионными перестройками. Как правило, во всех случаях преобладают парацентрические инверсии [Стегний, 1981; Clauton et al., Coluzzi et al., 1979; Stone et al., 1960; Wasserman, 1963].
Возникновение инверсии как хромосомной мутации сопряжено с такими очевидными преобразованиями в кариотипе, как перевертывание блока хромосомы на 180° с соответствующим изменением порядка генов, включенных в инверсию, и подавление рекомбинации аллелей внутри инвертированного района у гетерозигот по инверсии. По-видимому, двойственное значение инверсий породило альтернативные гипотезы относительно их адаптивной роли. Согласно одной из них, запирание кроссинговера между участками с инверсионным и исходным порядком обеспечивает сохранность двух коадаптированных блоков аллелей. Вторая гипотеза оценивает инверсию как регуляторную мутацию, обеспечивающую изменение функциональной активности генов в связи с изменением их взаиморасположения в хромосоме и геноме в целом.
Считается, что впервые эта гипотеза была высказана Майром [Мауr, 1945] и поддержана Добржанским [Dobzhansky, 1948]. Однако Уайт [White, 1954] отмечает, что еще в 1937 г. Дарлингтон предположил, что инверсия, содержащая несколько полезных генов (аллелей), будет путем подавления кроссинговера препятствовать распаду комбинации. Прежде чем рассмотреть примеры "за" и "против" этой популярной в настоящее время гипотезы, необходимо подчеркнуть, что в своей основе она предполагает строгое сцепление (или абсолютную ассоциацию) альтернативных инверсионных последовательностей с определенными аллельными вариантами генных локусов.
Благодаря электрофоретическому анализу гено-ферментных систем появились многочисленные работы по выявлению ассоциативных связей аллоферментов с инверсиями. Нам известен лишь один пример жесткого сцепления локуса щелочной фосфатазы (АРН-2) с инверсией у Drosophila pavani [Nair, Brncic, 1971]. У этого вида локус АРН-2 имеет четыре аллеля (обозначаемых в соответствии с электрофоретической подвижностью цифрами 2, 3, 4, 5). Со стандартной последовательностью хромосомы 4R в 100 % случаев связан аллель 2, а с инвертированной - аллели 4 и 5.
Другие данные по сцеплению аллелей с инверсиями говорят лишь о некоторой статистически значимой ассоциации, а во многих случаях - об отсутствии какой-либо связи. Пракаш и Левонтин [Prakash, Lewontin, 1968, 1971] отмечают как жесткую ассоциацию инверсий хромосомы 3 у Drosophila pseudoobscura с аллелями личиночных белков (Pt-10 и Pt-12) и амилазы, хотя, по существу, связи эти относительны и одни и те же аллели встречаются в различных инверсионных последовательностях. Так, аллель 1,20 локуса Pt-12 отмечен в последовательности ST лишь с частотой 80 %, он же характерен для порядков AR (5 %) и TL (5 %). Аналогичные ситуации наблюдаются и в других случаях [Prakash, 1974; Prakash, Levitan, 1973, 1974; Prakash, Merritt, 1972]. Характерна пространственно-временная нестабильность подобных ассоциаций. Так, у D.robusta и D.melanogaster отсутствуют корреляции между изменениями частоты инверсий и включенных в них аллоферментов в пространстве видовых ареалов [Prakash, 1973; Voelker et al., 1978 ]. Существенные временные флуктуации сцепления инверсий с аллелями отмечены в популяциях D.melanogaster, где в отдельные годы наблюдений ассоциации вообще исчезали [Mukai, Voelker, 1977; Mukai et al., 1974].
Таким образом, данные по взаимоотношениям инверсий с ферментными локусами показывают, что гипотетически постулируемая жесткая коадаптация аллелей в инверсионных блоках практически крайне редка и скорее является исключением, чем правилом. Попытки объяснить наличие идентичных аллелей в разных инверсионных порядках независимым мутированием вряд ли обоснованны в связи с крайне низкой частотой мутационных событий, не сопоставимой с реальными частотами аллельных вариантов в инверсионных последовательностях. Кроме того, нелогично было бы ожидать появления и накопления "чужих" аллелей в инверсионном блоке с коадаптированной аллельной системой. Отбор "отметал" бы подобные сбалансированные варианты. Также некорректно объяснять такую ситуацию "просачиванием" аллелей из альтернативных инверсионных вариантов в результате двойного кроссинговера. Теоретически это событие крайне маловероятно; например, для D.melanogaster в гетерозиготе по инверсии I(2L) оно по расчету составляет 0,00011% [Mukai, Voelker, 1977].
Дополнительным фактом, противоречащим концепции строгой коадаптации аллелей в инверсиях, является то, что у большинства известных полиморфных генов (в частности, кодирующих ферменты) в популяциях обнаруживается более чем по два аллеля, что осложняет "привязку" мультиаллельных систем к двум хромосомным вариантам.
Разумеется, все вышеизложенное не отрицает того, что инверсии предотвращают рекомбинацию в участках хромосом, ими блокируемых. Напротив, имеются данные, что инверсии, могут подавлять кроссинговер локусов, далеко отстоящих от них [Charlesworth et al., 1979; Mukai et al., 1974]. И по-видимому, нет особых оснований считать, что комбинации аллельных вариантов генов, включаемых в инверсию в момент ее возникновения, оказывают определяющее влияние на судьбу инверсии. Что касается значимого неравновесия по сцеплению между самими ферментными локусами, то оно встречается и в отсутствие инверсий [Charlesworth, Charlesworth, 1973], хотя также нестабильно в пространстве и времени и отражает либо эпистатические взаимодействия адаптивного характера, контролируемые естественным отбором [Franklin, Lewontin, 1970 ], либо в отсутствии с теорией нейтральных аллелей [Kimura, Otha, 1971] - действие случайного генетического дрейфа.
| Рис. 4.1. Географическое распространение видов и схема видообразовании в палеарктическом комплексе малярийных комаров (Anopheles miaculipennis).
На схемах видовых ареалов круги со штриховкой обозначают места наших выборок соответствующих видов; гомосеквентные виды в схеме видообразования обведены пунктирными линиями: черными стрелками показаны наиболее реальные и белыми (пунктирными) - наименее реальные пути видообразования |
Подобную ситуацию мы можем видеть и у ряда хорошо изученных групп Drosophila. В группе virilis филогенетически исходные виды D. virilis, D. novamexicana и D. ezoana хромосомно мономорфны, в то время как более молодые виды D. americana americana, D. a.texana, D. lacicola и особенно D. montana отличаются инверсионным полиморфизмом [Stone et al., I960]. В обширной группе repleta хромосомно мономорфными также являются стволовые виды: D. peninsularis (подгруппа mercatorum); D. mulleri, D. aldrichi, D.wheeleri (подгруппа mulleri); D. fasciola, D. coroica (подгруппа fasciola). Производные от этих видов - соответственно D. paranaensis, D. mercatorum, D. pachuca, D. desertorum, D. pictilis, D. maju - имеют в своих кариофондах полиморфные инверсии [Wasserman, 1963]. Аналогичные закономерности характерны и для гавайских дрозофил [Carson et а1., 1967].
Таким образом, исследование некоторых видовых комплексов двукрылых показывает, что в центральной (стволовой) части филогенетических древ находятся хромосомно мономорфные (часто гомосеквентные) виды, и это является серьезным аргументом против участия полиморфных инверсий в видообразовательных событиях. Детальный анализ точек инверсионных разрывов демонстрирует, что в подавляющем большинстве случаев локализация фиксированных (межвидовых) и флуктуирующих (внутривидовых) инверсий не совпадает. В палеарктическом комплексе Anopheles maculipennis отмечено 9 фиксированных и около 30 флуктуирующих инверсий, причем все они уникальны по локализации, хотя отдельные точки хромосомных разрывов в ряде случаев исключительно близки. В virilis-группе рода Drosophila среди 43 фиксированных и 49 флуктуирующих только одна инверсия (2f), отмеченная в гетерозиготе у D. ezoana, зафиксирована у D. montana [Stone et al., I960]. В repleta-группе рода Drosophila (99 фиксированных и 45 флуктуирующих инверсий) имеются лишь 3 полиморфные инверсии, зафиксированные у производных видов [Wasserman, 1963]. Эти данные свидетельствуют против закономерной фиксации полиморфных инверсий предкового вида у производных видов.